1700000195
大氧化事件一直被认为是地球生命历史的关键转折点,但是近年来,科学界对其意义的认识发生了剧变。新的解释对本书的中心论点非常关键。过去研究人员认为,对生命而言,氧气是关键的环境决定因素:氧气并不限定地球上可以演化出哪种生命,而是如同松开了闸门,容许演化产生更大的复杂度。例如,动物的生活方式是运动,它们追逐猎物或者被追逐,这显然需要很多能量。而有氧呼吸提供的能量,比其他形式的呼吸作用高出几乎一个数量级,⑤因而不难设想,如果没有氧气,动物就不可能存在。这条逻辑如此直白,很少有人兴起疑问,但这正是麻烦所在:它掩盖了更深入的问题。我们认为动物需要氧气(实际上并非总是如此),环境中也存在氧气,那么氧气就是共同点,演化生物学的课题就是动物或植物的行为和特征。一切好像都顺理成章。
1700000196
1700000197
传统的地球生命史观念,一直以这种观点为不假思索的默认基础。我们通常认为,氧气是有益的,有利于生命。然而实际上,在太初世界的生物化学机制中,氧气所处的地位恰好相反:它具有强氧化性,是生命的毒药。教科书上的描述是,随着大气中的氧含量上升,这种危险的气体成为整个微生物世界强大的选择压力。有些学者甚至推测,微生物在压力下发生了大规模灭绝——马古利斯称这一事件为“氧气大屠杀”。虽然根本找不到任何化石证据证明这种大规模灾难事件发生过,支持者却不以为意;他们为此找到的借口是微生物太小,事件发生的时间太过久远,证据早已湮没。氧气迫使单细胞生命形成新的生存关系:共生和内共生,细胞通过在内部或外部共享或交换代谢工具继续生存。几亿年间,生命的复杂度不断增加,细胞不仅学会了抵抗氧气,还学会了利用其化学活性:细胞演化出有氧呼吸,为自身提供更多的能量。这些大型、复杂、进行有氧呼吸的细胞把它们的DNA聚集在一个特化的结构内,形成了细胞核,并由此得名为“真核细胞”。我再提醒一次读者,以上是教科书上的标准理论。但我认为,它错了。
1700000198
1700000199
今天,我们周围所有的复杂生命,包括所有的植物、动物、藻类、真菌和原生生物(阿米巴原虫等大型单细胞生物)在内,都由真核细胞构成。真核生物在出现后的十亿年间,逐渐在生物圈取得统治地位,这个地质时期又被称为“无趣的十亿年”,因为化石证据稀缺。然而,研究人员在16亿~12亿年前的地质层中发现了单细胞生物的化石。它们像极了真核生物,其中一些的特征甚至非常符合现代生物分类标准,例如红藻类和真菌类。
1700000200
1700000201
接下去的7.5亿~6亿年前,地球又经历了一个全球气候动荡期,伴随着一连串雪球地球时期。此后不久,大气氧含量迅速上升至接近现代水平,最初的动物化石突然出现在化石记录中。最早的大型化石(直径可达1米)是神秘的埃迪卡拉生物群(又名文德纪生物),外形像对称的蕨状叶。大多数古生物学家认为它们是滤食性动物,也有一些学者认为它们不过是地衣。紧接着,绝大多数埃迪卡拉生物就在化石记录中消失了,灭绝和出现一样突兀。5.41亿年前的寒武纪初期,可以明确辨认的动物类群爆炸式地出现。对生物学家来说,这个日期就像1066年或者1492年对于历史学家那样意义非凡⑥。寒武纪动物的个头更大,能运动,长着复杂的眼和攻击性附肢;这些凶猛的捕食者,还有它们那些形状可怕、长有甲壳的猎物突然涌入演化的竞技场,牙尖爪利,物竞天择。
1700000202
1700000203
以上版本的地球生命史,究竟有多少内容是错的呢?表面上看,这一套似乎言之成理。但在我看来,其理论内涵首先就有误;随着知识的积累,我们会发现,很多细节也是错的。理论内涵的问题在于基因与环境的互动。对上述整个场景的描述都围绕着氧气进行,认为氧气是关键的环境变量,它松开了能量供应对基因创新的约束,从而促成遗传演化。大气氧含量明显上升了两次,分别是在24亿年前的大氧化事件时期,以及6亿年前漫长的前寒武纪最后阶段。氧气含量的每一次上升都解放了新的细胞结构和功能。大氧化事件同时带来了新的威胁和机遇,细胞通过一系列内共生,彼此交换、整合,逐渐积累起真核细胞的复杂度。而氧含量在寒武纪大爆发之前的第二次上升,彻底打破了细胞此前受到的物理束缚,就像魔术师的斗篷挥过,刹那间动物生命的无限可能喷涌而出。没人说是氧气本身驱动了这些改变;氧气只是让自然选择发挥作用的大环境完全改观。在这个不受束缚、海阔天空的新环境中,基因组自由扩张,它们携带的信息千变万化。生命欣欣向荣,以无穷无尽的形式充满所有的生态位。
1700000204
1700000205
这种演化史观基于辩证唯物主义,代表了20世纪中前期形成的新达尔文主义演化生物学观点。这一派认为,演化生物学中主要的对立互动因素是基因和环境,也可称为先天和后天。生物完全由基因决定,而它们的行为又完全取决于环境。除此之外,还能有什么呢?然而,真实世界的生物学并不只有基因和环境,还有细胞,以及约束它们物理结构的条件,而这些因素与基因和环境并没有直接关系。两种迥异的世界观会导致截然不同的预测。
1700000206
1700000207
先看第一种观念,即以基因和环境的互动来解释演化。早期地球缺乏氧气,这是最主要的环境约束。加入氧气这个变量后,生物演化就马上蓬勃发展。所有被氧气包围的生物都会受其影响,它们必须适应新的环境。有些细胞恰巧比其他细胞更适应有氧环境,因此可以繁衍生息下去;不适应者则自然灭绝。当然,地球上有很多不同的局部微环境,增加的氧气并没有充满整个世界,所以并没有形成单一的全球生态系统。氧气氧化各类矿物质,进而溶解于海洋,使各种无氧环境也变得更加丰富,硝酸盐、亚硝酸盐、硫酸盐和亚硫酸盐等各种物质供应都相应增加。细胞可以用这些物质来代替氧气进行呼吸作用,因此在无氧环境中,无氧呼吸作用也很旺盛。所有这些影响加在一起,造就了一个拥有多样生命形式的全新世界。
1700000208
1700000209
想象在某个环境中有一群随机混合的细胞。有些细胞,如阿米巴原虫,通过吞噬其他细胞生活,这种行为名为吞噬作用;有些细胞能进行光合作用;另一些细胞,比如真菌,能通过渗透营养(osmotrophy)在外部消化食物。假定细胞结构上没有不可逾越的约束条件,我们会推测这些不同种类的细胞源自不同的细菌祖先。某个祖先细菌可能会进行某种原始的吞噬作用,另一个擅于简单的渗透营养,另一个则会进行光合作用。日积月累,它们的后代越来越特化,越来越适应某种特定的环境和代谢方式。
1700000210
1700000211
用更专业的术语来描述就是,如果氧气水平上升能让新的生命形式蓬勃发展,我们就会观察到多系辐射演化(polyphyletic radiation),即在同一时段的同一环境中,没有亲缘关系的各种单细胞或多细胞生物(来自不同的门)迅速适应,辐射出新的种系,填充空白的生态位。我们确实在某些情况下观察到了这种演化模式。例如在寒武纪大爆发中,海绵、棘皮动物、节肢动物和蠕虫等几十个不同种系的动物辐射演化。伴随它们出现的是藻类、真菌和原生生物(例如纤毛虫)的辐射演化。生态系统变得更加复杂,因此又驱动了更进一步的变化。无论究竟是不是氧气水平的上升触发了寒武纪大爆发,学界的基本共识是,环境的变化确实重塑了自然选择的模式。重要的变化发生了,世界从此改变。
1700000212
1700000213
如果细胞结构上的限制是主导因素,那么我们观察到的现象就不会符合这种模式。如果结构限制不变,无论环境怎么变化,生物的变化都很有限。我们会看到长时期的演化停滞,对环境变化无动于衷,偶尔会出现单源辐射演化(monophyletic radiation):在非常罕见的情况下,某个种群克服了自身特有的结构限制。那么,只有这种生物会产生辐射演化(而且很可能会推迟,直到历经合适的环境变化后才会发生),其后代会填充生态位空白。当然,这也是我们观察到的实际情况。我们在寒武纪大爆发中看到了不同动物种群的辐射演化,但它们并非起源于不同的动物。所有的动物种群都拥有共同的祖先,植物也一样。对于复杂的多细胞生物来说,其特定的种系和身体构建是一项难度惊人的工程:构建程序极其精密复杂,对个体细胞的命运施加严格的控制,因此产生了对细胞结构的限制。在全局层面,这种限制相对较为宽松,各种生物的多细胞构建程序既有相同成分,又有细节差异。在所有生物中,大约有30种不同起源的多细胞构建模板,包括藻类(海藻)、真菌和黏菌。然而,在一种特殊情况下,细胞的结构限制成了压倒一切的主导因素:从细菌到真核细胞(大型复杂细胞)的起源。这恰好发生在大氧化事件之后。
1700000214
1700000215
生物学核心的黑洞
1700000216
1700000217
如果复杂的真核细胞演化确实源自大气中氧含量的上升,那么我们应该发现多系辐射演化,即不同种群的细菌独立发展出不同的复杂细胞种类。我们应该观察到,光合作用细菌演化为更大、更复杂的藻类,渗透营养细菌演化为真菌,运动捕食细胞演化为噬菌生物,以此类推。这种迈向更高复杂度的演化,可以通过标准的基因变异、基因交换和自然选择等方式进行,也可以通过细胞融合与摄入等方式进行,正如马古利斯在她的系列内共生理论中设想的那样发展。如果细胞结构方面不存在根本的限制条件,无论具体采用哪种演化方式,氧气的增长总会催生出更高的生物复杂度。氧气会突破所有细胞的能量限制,让所有种类的细菌独立演化,造成多源辐射演化。但是,我们观察到的实际情况并非如此。
1700000218
1700000219
容我通过更多细节展开,因为其中的推理过程至关重要。如果复杂细胞是通过“标准”的自然选择过程演化而来的,即基因变异导致多样化,自然选择从中遴选适者,那么,我们观察到的各种细胞内部结构,就会和细胞的外观一样迥然各异。真核细胞的大小和形状差异极大,从巨大的叶状藻类细胞到纺锤状神经元,还有随意延展的阿米巴原虫。如果真核生物的不同种群因为亿万年来在不同环境中适应不同的生活方式而发展出各自的复杂度,那么,如此久远的演化历程会反映为完全不同的内部结构。但是,请仔细观察真核细胞的内部细节。我们会发现,所有的真核细胞内部,都由基本相同的部件构成。在电子显微镜下,绝大多数人都无法分辨一个植物细胞、一个肾脏细胞和家门口池塘里的一个原生生物细胞:它们看起来全都一样。请看图3。如果大气氧含量的升高移除了对复杂度的限制,“标准”自然选择演化理论的预期是,不同种群适应不同的生境,这将导致多系辐射演化。但事实并非如此。
1700000220
1700000221

1700000222
1700000223
1700000224
图3 真核细胞的复杂度
1700000225
1700000226
四种不同的真核细胞表现出相似的形态复杂度。小图A是一个动物细胞(血浆细胞),有大型的中央细胞核(N)和线粒体(M),层层叠叠的内膜(内质网,ER)镶嵌着核糖体。小图B是裸藻属的单细胞藻类生物眼虫藻(Euglena),常见于池塘,能看见其中央细胞核(N)、叶绿体(C)和线粒体(M)。小图C是被细胞壁包被的植物细胞,有液泡(V)、叶绿体(C)、细胞核(N)和线粒体(M)。小图D是壶菌(一种真菌)的游动孢子,它已导致150种蛙类的灭绝。(N)细胞核,(M)线粒体,(F)鞭毛,(G)是功能未知的伽马小体。
1700000227
1700000228
马古利斯自20世纪60年代后期开始发表自己的理论,她认为以上说法是错误的:真核细胞的演化不是通过标准的自然选择,而是通过系列内共生实现的。某些细胞迫于环境压力紧密地合作共生,最终,其中一些进入另一些体内生活。这种理论的源头,可以追溯到20世纪初的理查德·阿尔特曼(Richard Altmann)、康斯坦丁·梅利什科夫斯基(Konstantin Mereschkowski)、乔治·波蒂捷(George Portier)和伊万·瓦林(Ivan Wallin)等科学家。他们认为,所有的复杂细胞都起源于简单细胞之间的共生。他们的理论没有被人遗忘,但遭到了无情的嘲笑:“纯属异想天开,这在正经的生物学讨论中不值一提。”直到60年代分子生物学取得革命性进展,马古利斯的理论才获得了更坚实的基础,虽然仍存在很大的争议。我们现在知道,真核细胞至少有两个部件来源于细菌内共生:线粒体(复杂细胞中的能量转换器)起源于α-变形菌,叶绿体(植物细胞的光合作用机器)起源于蓝细菌。真核细胞中其余所有的特化细胞器,都曾在不同的阶段被看作内共生体,包括细胞核、纤毛和鞭毛(纤细弯曲的构造,以节律性的旋转来驱动细胞运动),以及过氧化物酶体(进行毒性物质代谢的细胞器)。因此,系列内共生理论认为,真核细胞本质上是由多种细菌整合而成的,在大氧化事件之后的几亿年间,细菌的合作共生慢慢发展成一种“公共企业”,也就是真核生物。
1700000229
1700000230
这真是史诗一般的瑰丽观念,但是系列内共生理论的间接推论与标准的自然选择理论殊途同归。如果它成立,我们应该观察到细胞内部结构的多源性,其差异程度与细胞外观之间的差异相当。任何一个内共生实例都发生在一个特定的环境中,细胞共生总是要依赖于某种适应这个环境的代谢交换。那么,既然有那么多不同的环境,就应该有不同类型的细胞间代谢互动。如果这些细胞后来特化为复杂真核细胞的细胞器,那么内共生假说应该有以下推论:一些真核细胞有一套特定的部件,另一些有不同的一套。我们还应该在死水淤泥等各种生态角落中发现各种中间型,以及各种无亲缘关系的变异种类。2011年,马古利斯不幸因为中风而过早离开人世。直至去世前,她一直坚持自己的信念,认为真核生物就是由内共生形成的繁复拼图。对她来说,内共生是一种基本的生活方式,是很少有人探索的“雌性”演化途径,其中的合作因素(她的叫法是“联网”)超越了令人反感的“雄性”竞争因素,即捕猎者与猎物的关系。然而,马古利斯迷醉于“真正”的生命细胞,却忽视了另一个枯燥但极其重要的学术领域:种系发生学。科系发生学会对基因序列和整个基因组进行定量研究,能够告诉我们不同的真核生物之间究竟存在着怎样的亲缘关系。通过这方面研究而建立起来的理论,与马古利斯的假说大异其趣,而且更有说服力。
1700000231
1700000232
这项研究基于一大类(超过1,000种)简单的单细胞真核生物,它们的共同特征是没有线粒体。科学家曾经以为,这些生命是从细菌到复杂真核生物的演化中“缺失的环节”,恰好是系列内共生理论曾经预言过的中间型。这类生物包括梨形鞭毛虫(Giardia),一种危害严重的肠道寄生虫,科普作家埃德·扬(Ed Yong)曾把它比作“一滴邪恶的眼泪”。它的危害与其造型相称,会导致恶性腹泻。它的细胞拥有不止一个而是两个细胞核,所以毫无疑问是真核细胞。但它缺少真核细胞的其他结构特征,尤其是没有线粒体。20世纪80年代中期,标新立异的生物学家汤姆·卡瓦利耶-史密斯(Tom Cavalier-Smith)提出,梨形鞭毛虫和其他一些较为简单的真核生物,很可能是自真核细胞演化早期残存至今的生命,早在真核细胞获取线粒体之前。卡瓦利耶-史密斯同意线粒体源自细菌内共生的假说,但他对马古利斯后来的系列内共生理论没有多少耐心。直到现在他都认为最早的真核生物是原始的噬菌生物,类似于现代的阿米巴原虫,通过吞噬其他细胞生存。他还认为,最早获得线粒体的细胞已经有细胞核,也有动态的细胞“内骨架”,能够变形、移动,另外还有进行胞内物质运输的蛋白结构,以及专门用于消化食物的内部分区,等等。获取线粒体当然很有好处,因为线粒体大大提升了这些原始细胞的能量水平。但是,对一辆汽车进行动力改装,并不会改变汽车的基本架构:你用到的仍然是发动机、变速箱、刹车等一切基本的汽车部件。动力改装只是增加了汽车的输出功率。卡瓦利耶-史密斯的原始噬菌细胞假说与此同理,除了线粒体,其他所有部件都已就位,线粒体只是为细胞提供了更多能量。截至目前,这是关于真核细胞起源最正统的理论。
1700000233
1700000234
1700000235
1700000236
1700000237
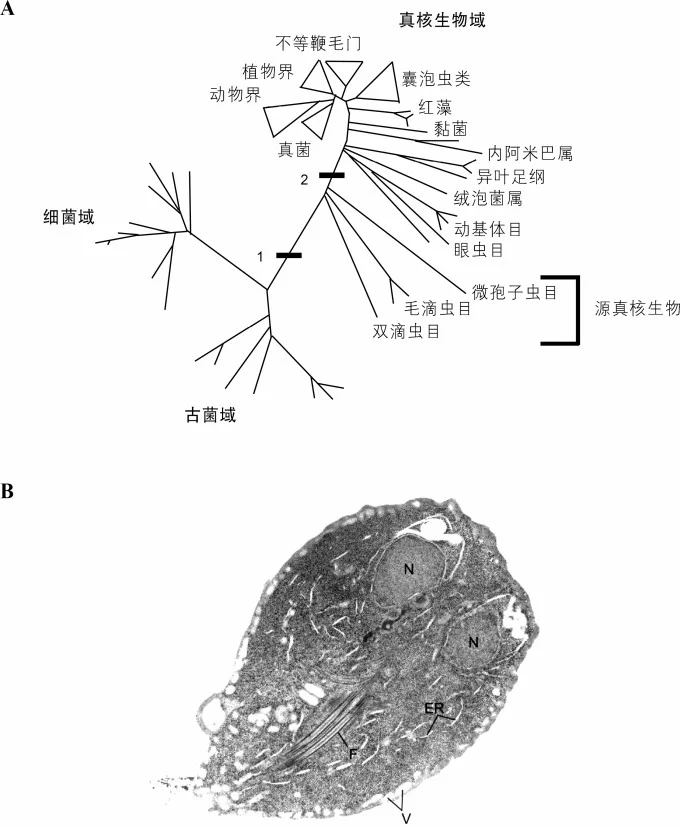
图4 源真核生物,曾经被误认为演化过程中“缺失的环节”
1700000238
1700000239
小图A是早期不正确的生命树图,是基于核糖体RNA研究绘制的。图中分为三个域(生物学中最大的分类单位):细菌域、古菌域和真核生物域。图中标记条1表示细胞核的早期演化,标记条2表示后来获取了线粒体。两条标记条之间发出的三个分支组成了源真核生物,理论上还没有获取线粒体的原始真核生命,例如梨形鞭毛虫(小图B)。我们现在已经明确,所谓的源真核生物根本不是原始的真核生命,而是源于拥有线粒体的更加复杂的祖先。它们正确的分支点应该在真核生物分支的内部。(N=细胞核,ER=内质网,V=液泡,F=鞭毛)
1700000240
1700000241
卡瓦利耶-史密斯把这些早期的真核生物命名为“源真核生物”(archezoa,意为“古代动物”),以强调它们的古老程度。其中有几种是致病的寄生虫,因此它们的生物化学细节和基因组引起了医学研究者的兴趣,随之带来了研究经费。所以,我们现在对它们了解得非常深入。过去的20年间,我们通过它们的基因组序列和生物化学细节得出了结论:源真核生物中没有一种是真正意义上的缺失环节,它们也不是真正的演化中间型。恰恰相反,它们全部起源于更复杂的真核生物,这些祖先曾经拥有所有的真核细胞特征,包括线粒体。源真核生物(注意这一术语本身的误导性)为了适应它们所处的简单生态环境,失去了其祖先曾经拥有的复杂度。它们全都保留着氢酶体或者纺锤剩体这样的结构部件,实际上是线粒体退化的产物。这些部件虽然同样有双层膜结构,但从外观看并不像线粒体,这才造成了“源真核生物没有线粒体”的误解。分子生物学和种系发生学的研究数据也证明,氢酶体和纺锤剩体起源于线粒体,而非源于某种细菌的内共生(根据马古利斯的理论)。所以,所有真核生物都有线粒体,虽然形式上可能会有所不同。我们可以推论:所有真核生物的最后共同祖先已经拥有线粒体,正如马丁于1998年预言的那样(详见绪论)。“所有真核生物都有线粒体”,乍一看来好像并不起眼。然而,随着对微生物世界基因组测序的广泛开展,这一认知彻底颠覆了科学界对真核生物演化的理解。
1700000242
1700000243
我们现在知道,所有的真核生物都有一个共同的祖先。这个“共同祖先”的生物学定义意味着,在40亿年的地球历史中它只出现过一次。让我再次强调这个概念的重要意义:所有的植物、动物、藻类、真菌和原生生物都有共同的祖先——真核生物是单源性的!也就是说,植物、动物和真菌并非分别从不同的细菌演化而来,恰好相反:一个形态复杂的真核细胞种群在某个特定历史场合出现,而所有的植物、动物、藻类和真菌都演化自这个始祖种群。共同祖先,概念上就必须是单一的实体——不是指一个单独的细胞,而是一个单独的种群,其中所有的细胞本质上完全相同。这个概念本身,并不意味着复杂细胞的起源是演化中的稀有事件。理论上,复杂细胞可能出现过很多次,但只有一个种群生存下来,后代繁衍至今,而其他的都早已灭绝。我认为实际情况并非如此,但是,首先我们必须更详细地考虑真核生物的特征。
1700000244