1700000224
图3 真核细胞的复杂度
1700000225
1700000226
四种不同的真核细胞表现出相似的形态复杂度。小图A是一个动物细胞(血浆细胞),有大型的中央细胞核(N)和线粒体(M),层层叠叠的内膜(内质网,ER)镶嵌着核糖体。小图B是裸藻属的单细胞藻类生物眼虫藻(Euglena),常见于池塘,能看见其中央细胞核(N)、叶绿体(C)和线粒体(M)。小图C是被细胞壁包被的植物细胞,有液泡(V)、叶绿体(C)、细胞核(N)和线粒体(M)。小图D是壶菌(一种真菌)的游动孢子,它已导致150种蛙类的灭绝。(N)细胞核,(M)线粒体,(F)鞭毛,(G)是功能未知的伽马小体。
1700000227
1700000228
马古利斯自20世纪60年代后期开始发表自己的理论,她认为以上说法是错误的:真核细胞的演化不是通过标准的自然选择,而是通过系列内共生实现的。某些细胞迫于环境压力紧密地合作共生,最终,其中一些进入另一些体内生活。这种理论的源头,可以追溯到20世纪初的理查德·阿尔特曼(Richard Altmann)、康斯坦丁·梅利什科夫斯基(Konstantin Mereschkowski)、乔治·波蒂捷(George Portier)和伊万·瓦林(Ivan Wallin)等科学家。他们认为,所有的复杂细胞都起源于简单细胞之间的共生。他们的理论没有被人遗忘,但遭到了无情的嘲笑:“纯属异想天开,这在正经的生物学讨论中不值一提。”直到60年代分子生物学取得革命性进展,马古利斯的理论才获得了更坚实的基础,虽然仍存在很大的争议。我们现在知道,真核细胞至少有两个部件来源于细菌内共生:线粒体(复杂细胞中的能量转换器)起源于α-变形菌,叶绿体(植物细胞的光合作用机器)起源于蓝细菌。真核细胞中其余所有的特化细胞器,都曾在不同的阶段被看作内共生体,包括细胞核、纤毛和鞭毛(纤细弯曲的构造,以节律性的旋转来驱动细胞运动),以及过氧化物酶体(进行毒性物质代谢的细胞器)。因此,系列内共生理论认为,真核细胞本质上是由多种细菌整合而成的,在大氧化事件之后的几亿年间,细菌的合作共生慢慢发展成一种“公共企业”,也就是真核生物。
1700000229
1700000230
这真是史诗一般的瑰丽观念,但是系列内共生理论的间接推论与标准的自然选择理论殊途同归。如果它成立,我们应该观察到细胞内部结构的多源性,其差异程度与细胞外观之间的差异相当。任何一个内共生实例都发生在一个特定的环境中,细胞共生总是要依赖于某种适应这个环境的代谢交换。那么,既然有那么多不同的环境,就应该有不同类型的细胞间代谢互动。如果这些细胞后来特化为复杂真核细胞的细胞器,那么内共生假说应该有以下推论:一些真核细胞有一套特定的部件,另一些有不同的一套。我们还应该在死水淤泥等各种生态角落中发现各种中间型,以及各种无亲缘关系的变异种类。2011年,马古利斯不幸因为中风而过早离开人世。直至去世前,她一直坚持自己的信念,认为真核生物就是由内共生形成的繁复拼图。对她来说,内共生是一种基本的生活方式,是很少有人探索的“雌性”演化途径,其中的合作因素(她的叫法是“联网”)超越了令人反感的“雄性”竞争因素,即捕猎者与猎物的关系。然而,马古利斯迷醉于“真正”的生命细胞,却忽视了另一个枯燥但极其重要的学术领域:种系发生学。科系发生学会对基因序列和整个基因组进行定量研究,能够告诉我们不同的真核生物之间究竟存在着怎样的亲缘关系。通过这方面研究而建立起来的理论,与马古利斯的假说大异其趣,而且更有说服力。
1700000231
1700000232
这项研究基于一大类(超过1,000种)简单的单细胞真核生物,它们的共同特征是没有线粒体。科学家曾经以为,这些生命是从细菌到复杂真核生物的演化中“缺失的环节”,恰好是系列内共生理论曾经预言过的中间型。这类生物包括梨形鞭毛虫(Giardia),一种危害严重的肠道寄生虫,科普作家埃德·扬(Ed Yong)曾把它比作“一滴邪恶的眼泪”。它的危害与其造型相称,会导致恶性腹泻。它的细胞拥有不止一个而是两个细胞核,所以毫无疑问是真核细胞。但它缺少真核细胞的其他结构特征,尤其是没有线粒体。20世纪80年代中期,标新立异的生物学家汤姆·卡瓦利耶-史密斯(Tom Cavalier-Smith)提出,梨形鞭毛虫和其他一些较为简单的真核生物,很可能是自真核细胞演化早期残存至今的生命,早在真核细胞获取线粒体之前。卡瓦利耶-史密斯同意线粒体源自细菌内共生的假说,但他对马古利斯后来的系列内共生理论没有多少耐心。直到现在他都认为最早的真核生物是原始的噬菌生物,类似于现代的阿米巴原虫,通过吞噬其他细胞生存。他还认为,最早获得线粒体的细胞已经有细胞核,也有动态的细胞“内骨架”,能够变形、移动,另外还有进行胞内物质运输的蛋白结构,以及专门用于消化食物的内部分区,等等。获取线粒体当然很有好处,因为线粒体大大提升了这些原始细胞的能量水平。但是,对一辆汽车进行动力改装,并不会改变汽车的基本架构:你用到的仍然是发动机、变速箱、刹车等一切基本的汽车部件。动力改装只是增加了汽车的输出功率。卡瓦利耶-史密斯的原始噬菌细胞假说与此同理,除了线粒体,其他所有部件都已就位,线粒体只是为细胞提供了更多能量。截至目前,这是关于真核细胞起源最正统的理论。
1700000233
1700000234
1700000235
1700000236
1700000237
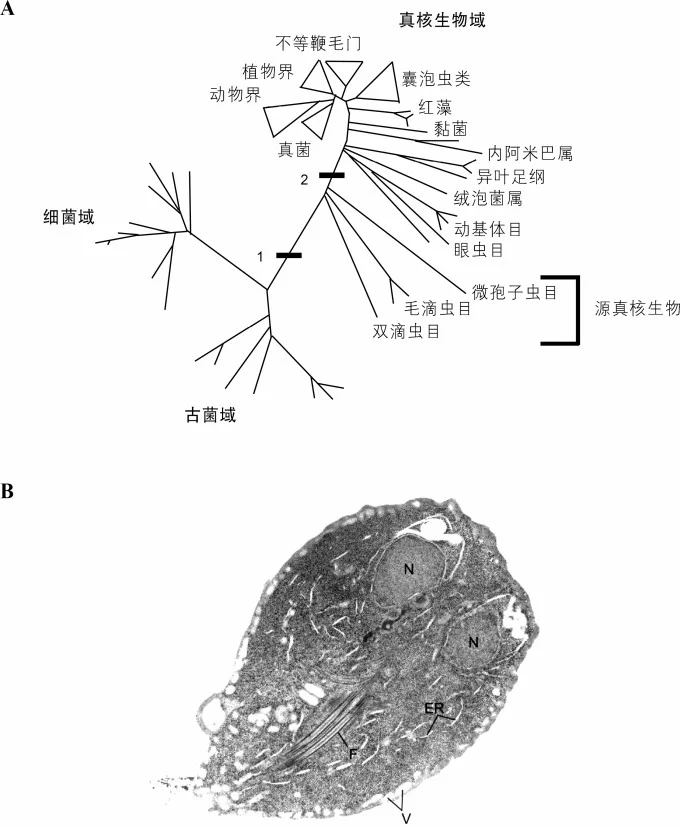
图4 源真核生物,曾经被误认为演化过程中“缺失的环节”
1700000238
1700000239
小图A是早期不正确的生命树图,是基于核糖体RNA研究绘制的。图中分为三个域(生物学中最大的分类单位):细菌域、古菌域和真核生物域。图中标记条1表示细胞核的早期演化,标记条2表示后来获取了线粒体。两条标记条之间发出的三个分支组成了源真核生物,理论上还没有获取线粒体的原始真核生命,例如梨形鞭毛虫(小图B)。我们现在已经明确,所谓的源真核生物根本不是原始的真核生命,而是源于拥有线粒体的更加复杂的祖先。它们正确的分支点应该在真核生物分支的内部。(N=细胞核,ER=内质网,V=液泡,F=鞭毛)
1700000240
1700000241
卡瓦利耶-史密斯把这些早期的真核生物命名为“源真核生物”(archezoa,意为“古代动物”),以强调它们的古老程度。其中有几种是致病的寄生虫,因此它们的生物化学细节和基因组引起了医学研究者的兴趣,随之带来了研究经费。所以,我们现在对它们了解得非常深入。过去的20年间,我们通过它们的基因组序列和生物化学细节得出了结论:源真核生物中没有一种是真正意义上的缺失环节,它们也不是真正的演化中间型。恰恰相反,它们全部起源于更复杂的真核生物,这些祖先曾经拥有所有的真核细胞特征,包括线粒体。源真核生物(注意这一术语本身的误导性)为了适应它们所处的简单生态环境,失去了其祖先曾经拥有的复杂度。它们全都保留着氢酶体或者纺锤剩体这样的结构部件,实际上是线粒体退化的产物。这些部件虽然同样有双层膜结构,但从外观看并不像线粒体,这才造成了“源真核生物没有线粒体”的误解。分子生物学和种系发生学的研究数据也证明,氢酶体和纺锤剩体起源于线粒体,而非源于某种细菌的内共生(根据马古利斯的理论)。所以,所有真核生物都有线粒体,虽然形式上可能会有所不同。我们可以推论:所有真核生物的最后共同祖先已经拥有线粒体,正如马丁于1998年预言的那样(详见绪论)。“所有真核生物都有线粒体”,乍一看来好像并不起眼。然而,随着对微生物世界基因组测序的广泛开展,这一认知彻底颠覆了科学界对真核生物演化的理解。
1700000242
1700000243
我们现在知道,所有的真核生物都有一个共同的祖先。这个“共同祖先”的生物学定义意味着,在40亿年的地球历史中它只出现过一次。让我再次强调这个概念的重要意义:所有的植物、动物、藻类、真菌和原生生物都有共同的祖先——真核生物是单源性的!也就是说,植物、动物和真菌并非分别从不同的细菌演化而来,恰好相反:一个形态复杂的真核细胞种群在某个特定历史场合出现,而所有的植物、动物、藻类和真菌都演化自这个始祖种群。共同祖先,概念上就必须是单一的实体——不是指一个单独的细胞,而是一个单独的种群,其中所有的细胞本质上完全相同。这个概念本身,并不意味着复杂细胞的起源是演化中的稀有事件。理论上,复杂细胞可能出现过很多次,但只有一个种群生存下来,后代繁衍至今,而其他的都早已灭绝。我认为实际情况并非如此,但是,首先我们必须更详细地考虑真核生物的特征。
1700000244
1700000245
所有真核生物的共同祖先很快繁衍出了五个细胞形态各异的生物“超类群”(supergroups),其中大多数,甚至对接受过正统学术训练的生物学家都很陌生。这些超类群有诸如“单鞭毛界”(unikonts,包括动物和真菌)、“古虫界”(excavates)、“囊泡藻界”(chromalveolates)和“植物界”(plantae,包括陆地植物和藻类)等古怪的名字。名字不重要,重要的是两个细节。第一,每个超类群内部的基因多态性,远远超出各个超类群祖先之间的基因差异(图5)。这标志着早期的爆发性辐射演化,准确地说是单源辐射演化,意味着生命体从某种结构限制中解脱出来。第二,真核生物的共同祖先已经是非常复杂的细胞。通过比较各个超类群之间的特征,我们能归纳出共同祖先可能具有的特征。每个超类群中每个物种的共同特征,有理由认为继承自共同祖先;只有一两个超类群中才有的特征,则可以认为是后续演化出来的。叶绿体是后一种情况的典型例子:它仅见于植物界和囊泡藻界,是广为人知的内共生演化结果。所以,真核生物的共同祖先没有叶绿体。
1700000246
1700000247

1700000248
1700000249
1700000250
图5 真核生物的超类群
1700000251
1700000252
库宁于2010年基于几千个基因制作的真核生物树,显示5个超类群。图中的数字表示超类群和真核生物最后共同祖先(LUCA)共有的基因个数。每个超类群都独立获得或丢失了其他很多基因。单细胞原生生物的差别最大。注意每个超类群的内部差异远大于各群之间的祖先差异,这反映了爆炸性的辐射演化。我喜欢中间那个富有深意的“黑洞”。真核生物的共同祖先已经演化出了所有的真核生物共同特征,但是种系发生学不能告诉我们这些特征是怎么从细菌或古菌演化而来的。这是一个位于演化中心的黑洞。
1700000253
1700000254
那么,根据种系发生学的研究,哪些特征才是共同祖先固有的呢?答案令人惊讶:几乎所有其他特征都是。让我们来看一下几种典型的细胞成分。我们已知共同祖先有细胞核,其中贮藏着DNA。细胞核的结构非常复杂,所有真核生物都保留了下来。细胞核被双层膜包裹,或者说是很多看起来像双层膜的扁囊状膜结构,并续接至细胞质中的内质网膜。核膜上镶嵌着精细的蛋白质核孔,具有弹性内衬支撑结构。细胞核内部的其他结构,比如核仁,同样在所有真核生物中保留。值得强调的是,这些复杂结构中有十多种发挥核心功能的蛋白质,都存在于所有的超类群中;包裹着DNA的各种组蛋白也是如此。所有真核生物的DNA都是线性染色体,两头有端粒,其功能类似于鞋带上的顶盖,可以防止染色体两端“磨损”。真核细胞的基因是所谓的“碎片基因”:编码蛋白质的基因被分成很多个小片段(外显子),散布在长长的非编码区域(内含子)之间。在基因转译成蛋白质之前,内含子会被剪切出去,而这种剪切的生化机制在所有真核生物中都一样。内含子所处的位置,很多时候也都保留下来:在各种真核生物的同一个基因上,同一个序列位置会插入相似的内含子。
1700000255
1700000256
细胞核之外的情况也差不多。除了简单的源真核生物(后来发现按照正确的分类,它们广泛分布在五个超类群中,再次证明了它们各自独立演化的事实,只是都失去了早先的复杂度而已),所有的真核生物都有基本相同的细胞结构:全都有复杂的内膜结构,例如内质网和高尔基体,后者的功能是包装和向外运输蛋白质;全都有动态的细胞骨架,能够进行各种变形,满足各类功能需求;全都有马达蛋白,作用是沿着细胞骨架的微管或微丝来回运送物质;全都有线粒体、溶酶体、过氧化物酶体、胞内和跨膜运输机制,以及相同的生物化学信号系统。真核生物的其他共同特征不胜枚举:所有真核细胞都进行有丝分裂,同源染色体在一个由微管构成的纺锤体上分离,由同一组酶控制。所有的真核生物都进行有性生殖,生命周期中都经历减数分裂,以形成配子(例如精子和卵子);配子再结合成合子,开始下一周期。极少数失去了有性生殖特征的真核生物,通常很快就会灭绝(在演化生物学的时间尺度下,“很快”一般指几百万年)。
1700000257
1700000258
从前我们通过显微镜观察细胞结构就已经了解了上述的大多数情况,然而新时代的种系发生学研究澄清了两方面的问题。首先,结构上的相似性并非仅限于表面,而是源于相似的基因序列细节,由亿万个DNA字母组成。这种数据的积累,使我们能够通过分支树的形式,以空前的精确度计算生物的种系和亲缘关系。其次,高通量基因测序技术的出现,使自然界的取样研究不再依赖细胞培养和制作显微镜切片等烦冗的传统方法。霰弹枪定序法(又称鸟枪法)的速度和精度才是当今技术的极限代表。我们发现了几种意料之外的真核生物类型,包括真核嗜极生物,它能够利用高浓度的毒性金属进行代谢或者在高温下生活;还有尺寸极小但结构完整的单细胞“微型真核生物”,大小与细菌类似,但仍然拥有简化的细胞核和小型线粒体。我们如今对真核生物的多样性有了更清楚的认识。所有这些新发现的真核生物,都能被很清楚地归类到现存五大超类群中,它们并没有开辟出新的种系大类。如此高度的多样性反映了一条铁一般的事实:真核生物实在是太相似了。我们没有找到任何演化中间型或者非亲缘变体,系列内共生理论预言我们应该找到很多,所以它不正确。
1700000259
1700000260
这又带来了新的问题。种系发生学的惊人成功和生物学研究的信息化,很容易让我们忽视它们的局限性。用种系发生学方法研究真核生物起源,会受困于“事件视界”(event horizon)⑦,止步于“黑洞”边缘。所有的基因组都可以回溯到真核生物的最后共同祖先,而它几乎具备所有的真核生物特征。那么,所有这些部件又从何而来呢?真核生物共同祖先的出现,就像雅典娜从宙斯的头颅中跳出来,一出生就全副武装。⑧共同祖先出现之前,这些特征(也就是全部的特征)是怎么形成的?我们对这个问题的理解还极为浅薄。为什么会演化出细胞核?是如何演化的?性又是怎么来的?为什么几乎所有的真核生物都是两性的?繁复的细胞内膜结构源自何处?细胞骨架是怎么变得动态可塑的?为什么细胞的成熟分裂(即减数分裂)最终是为了让染色体数目减半,首先会使其倍增?为什么我们会衰老、会得癌症、会死去?种系发生学的研究方法虽然极尽精妙,却无法为生物学中这些核心问题提供多少答案。几乎所有涉及这些真核生物特征的基因(即编码真核生物“特征蛋白”的基因),都无法在原核生物中找到。反过来看,细菌也并未显示出任何向真核生物复杂特征演化的倾向。在原核生物的简单形态和真核生物复杂到令人不安的共同祖先之间,我们没有找到任何演化中间型(图6)。如此种种,都意味着复杂生命是从种系发生学的虚空中一跃而出。生物学的核心地带,存在着一个未知的黑洞。
1700000261
1700000262

1700000263
1700000264
1700000265
图6 生物学核心的黑洞
1700000266
1700000267
图中下方的细胞属于耐格里属(Naegleria),它的大小和复杂度都被认为近似于真核生物共同祖先。它具有细胞核(N)、内质网(ER)、高尔基体(Gl)、食物液泡(Fv)、吞噬体(Ps)和过氧化物酶体(P)。上方是一个比较复杂的细菌:浮霉菌(Planctomycetes),大致以同比例显示。此图并不表示下方的真核细胞是从浮霉菌演化来的(肯定不是),而是展示了一个复杂细菌和一个单细胞真核生物的相对比例和巨大差异。它们之间没有发现任何存活的中间种,所以无法提供演化的线索(用骷髅表示)。
1700000268
1700000269
复杂之路上的缺失环节
1700000270
1700000271
演化理论做过一项简单的预测:复杂特征产生自一系列微步演化,每一次微小的变化都会为生物提供相对于前一步的些许优势。自然选择保留最适应环境的特征,不适应的会遭到淘汰,所以自然选择会不断清除中间型。随着时间的积累,生物特征会达到适应的巅峰,所以我们会看到功能完美的眼睛,但无法发现眼睛演化历程中不够完美的中间形态。达尔文在《物种起源》中提出,自然选择机制本身就决定了中间型的缺失。在这个理论背景下,细菌和真核生物之间没有现存的中间型并不特别令人惊奇。真正出人意料的是,真核生物的特征并不像眼睛那样,在演化史上一再重现。
1700000272
1700000273
我们没有发现眼睛在演化史上的中间步骤,但确实发现了眼睛在生态系统中的连续分布。从一种古老蠕虫身上简陋的感光点开始,眼睛的独立演化发生了几十次。这正如自然选择学说预测的那样。在特定环境中,每一小步演化都会带来一点优势,具体是什么优势取决于具体的环境。生物在不同的环境下演化出形态各异的各种眼睛,差异演化使苍蝇的复眼与扇贝的镜面眼迥然不同,而收敛演化让人类和章鱼拥有非常相似的照相机式眼睛。我们能设想到的每种眼睛演化中间型,从小孔聚焦到可变焦透镜,都能在某种生物身上找到。我们甚至在一些单细胞的原生生物中发现了“微缩眼睛”,它具有类似“透镜”和“视网膜”的构造。简而言之,演化理论预言生物特征应该有多次起源,每次微步演化都会带来一些相对优势。理论上,这应该对每种生物特征都成立,也与我们平常的观察相符。动力飞行至少独立演化了6次,分别是蝙蝠、鸟类、翼龙和不同种类的昆虫;多细胞组织独立演化了大约30次,正如前面所述;不同形式的温血特征在好几类生物中独立出现,包括哺乳动物和鸟类,还包括某些特殊的鱼类、昆虫和植物;⑨甚至连大脑意识,似乎都是在哺乳动物和鸟类中分别独立出现的。正如眼睛的演化,我们观察到无数的形态差异,反映了各种形态在演化过程中经历的环境差异。物理上的限制固然存在,但并没有严苛到排除多次起源的程度。