1700189124
第二个重要的两足行走理论的说服力要大得多,部分的原因是因为它很简单。戴维斯加利福尼亚大学的人类学家彼得•罗德曼(Peter Rodman)和亨利•麦克亨利(Henry McHenry)提出假说,主张两足行走在环境条件改变时是有利的,因为它是一种更为有效的行动方式。当森林萎缩时,疏林生境的食物资源如果树,也变得太分散,使传统的猿难于有效地去利用。按照这个假说,最早的两足行走的猿只是在其行动方式上才是人。他们的手、上下颌和牙齿仍旧是像猿的,困为他们的食物并没有改变,只是他们获得食物的方式不同了。
1700189125
1700189126
对许多生物学家来说,这个假说最初被认为似乎是不大可能的。哈佛大学的研究者几年前曾显示两足行走不如四足行走有效(任何人都不会惊奇,带着一只狗或猫同时奔跑,令人难堪的是它们会比主人跑得快得多)。哈佛大学的研究者也曾比较了人类的两足行走和马与狗的四足行走之间的能耗率。可是罗德曼和麦克亨利指出,合适的比较应该是人和黑猩猩之间的比较。进行了这种比较后,证明人类的两足行走比黑猩猩的四足行走的效率要高得多。他们的结论是,以能量效益作为有利于两足行走的自然选择的力量是有道理的。
1700189127
1700189128
关于推动两足行走进化的因素,还有许多其他的意见,诸如为了监视猛兽,需要超出高草去观察;在大白天搜寻食物时,需要采取更为有效的冷却身体的姿势等等。在所有这些假说当中,我认为罗德曼和麦克亨利的假说最有说服力,因为它有牢固的生物学上的根据,符合最早的人类物种出现时的生态变化。如果这个假说是正确的,那就意味着,当我们发现最早的人类物种的时候,我们可能无法辨认他们。如果我们发现了一些骨头,如果是骨盆或是下肢骨,这都是两足行走运动的有效证据,我们就可以说它是“人”。但是如果我们发现的是头骨的某些部分,颌骨或是几个牙齿,它们可能看起来恰恰像是猿的。我们又怎样去辨别它们是属于两足行走的猿,还是属于传统意义上的猿呢?这对我们来说是一个挑战。
1700189129
1700189130
如果我们能访问700万年前的非洲,去观察最早人类的行为,那我们看到的模式,更像研究猿猴行为的灵长类学家所熟悉的模式,而不是研究人类行为的人类学家所熟悉的模式。最初人类的生活可能不像现代的狩猎—采集者那样由许多家庭集合成的游动群体,而是像稀树草原的狒狒那样。30个左右的人组成的群体,在一个大领域内以一种协调的方式搜寻食物,夜里回到安适的地方如悬崖下或树丛中去睡眠,成年女性和她们的子女是这个群体的大部分成员,只有少数几个成年男性。男性经常找寻交配的机会,其中占优势的个体成功的机会最多。未成年的和低级的雄性大部分时间是在群体的外围,往往自己找寻食物。群体中的各个个体像人那样地两足行走,但其他行为则类似稀树草原的灵长类。他们面临着700万年以来的进化,我们将看到这种进化类型是复杂的,也是很不确定的,因为自然选择是依赖某时某地的环境而进行,而没有一个长远的目标。智人最后的确是作为最初人类的后裔出现了,但是这并不一定是必然的。
1700189131
1700189132
1700189133
1700189134
1700189135
人类的起源 [:1700188756]
1700189136
人类的起源 第二章 一个拥挤的人科6
1700189137
1700189138
在南非和东非发现的最早人类化石的记录,即从大约400万年前到约100万年前,按我的计算,已有代表各种不同物种的至少1000个个体的标本,各个标本破碎的程度有所不同。较晚时期的化石则多得多。欧亚大陆发现的最早人类化石可能接近200万年(新大陆和澳洲有人居住是在晚得多的时期,大约分别为20000年和50000年以前)。因此可以合理他说,人类史前时代的大多数活动发生于非洲。关于这种活动,人类学家必须回答的问题有两方面:首先,什么物种于700万~200万年前之间在人类系统树上占有一席之地,他们是怎样生活的?其次,这些物种在进化上彼此之间有什么关系?换句话说,人类系统树是什么样子的?
1700189139
1700189140
我的人类学同事们在阐述这些问题时面临两项挑战。第一是达尔文所称的“地质学记录的极端不完整性”。在他的《物种起源》一书中,达尔文用整整一个章节的篇幅来谈论地质记录中令人沮丧的空缺内容。这些空缺是由于石化过程中变幻莫测的因素和骨骼化石后来暴露于外造成的结果。有利于骨骼的迅速掩埋和可能石化的条件是很罕见的。古代的沉积物可以经过侵蚀而暴露出来,例如一条河流切过沉积物。可是史前时代中哪些内容能以这种方式重新被揭开,则纯粹是机会问题,许多内容仍然会藏匿于地层中。例如,在东非这个最有希望的早期人类化石储藏所,只有很少的距今400万到800万年这段时期内的含化石沉积物。这是人类史前时代的关键时期,因为人科的起源发生在这个时期。虽然在400万年以后的时期中,我们所拥有的化石也远比我们想要有的少。
1700189141
1700189142
第二项挑战是被发现的大多数化石标本都是小的破片,一块头骨破片,一块颧骨,臂骨的一段和许多牙齿。由这样贫乏的证据来鉴定物种是很困难的,有时甚至是不可能的。由此而产生的不确定性使得许多不同的科学见解得以存在,差异既存在于物种的鉴定,也存在于各物种之间相互关系的辨别。人类学中被称为分类学和系统学的领域是争论最多的。我将避开许多争论的细节而集中于描述系统树的全面状况。
1700189143
1700189144
人类化石记录的知识在非洲缓慢地发展,这一发展开始于1924年,当时雷蒙德•达特宣布发现了著名的汤恩(Taung)小孩。这件标本包括一个小孩的不完整头骨,即部分颅骨,面骨、下颌骨和脑壳7,这个名称的由来是因为它是在南非的汤恩石灰岩采石场被发现的。虽然采石场堆积物的年代不可能精确地测定,但是科学的估计认为这个小孩生活在大约200万年前。
1700189145
1700189146
汤恩小孩的头骨有许多似猿的性状,例如小的脑子和向前突出的上、下颌骨,但是达特同时也还认识到它所具有的人类的性状:上、下颌骨不如猿向前突出,颊齿咬合面平,犬齿小。一个关键的证据是枕骨大孔的位置,枕骨大孔是头骨基底的开口,脊髓通过此孔进入脊柱。在猿类,此孔在颅底相对靠后的位置,而在人类则接近颅底中央。这种差别反映了人的两足行走的姿势,此时头是平衡于脊柱的顶端,与猿的姿势相反,猿的头向前倾。汤恩小孩的枕骨大孔在中央,表明此小孩是一个两足行走的猿。
1700189147
1700189148
虽然达特相信汤恩小孩属于人类,但是几乎过了四分之一世纪以后,人类学家们才接纳这个化石标本为人类的祖先而不只是一种古猿。反对非洲为人类进化地区的偏见和对如此像猿的一种生物可能是人类祖先这一想法普遍的反感结合起来,使达特和他的发现长期湮没无闻。在40年代后期,人类学家们认识到他们的错误。达特与苏格兰人罗伯特•布鲁姆结伴,从南非的4个山洞遗址[斯特克方丹(Sterkfontein),斯瓦特克朗(Swartkrans),克罗姆德莱(Kromdraai)及马卡潘斯盖特(Makapansgat)]中发现了大量早期人类化石。达特和布鲁姆遵循那时人类学的习惯,给他们发现的每一个化石命名了一个新的种名,因此不久之后看起来就似乎有了生活在300万年与100万年前之间的南非的一个包括不同种人类的动物园。
1700189149
1700189150
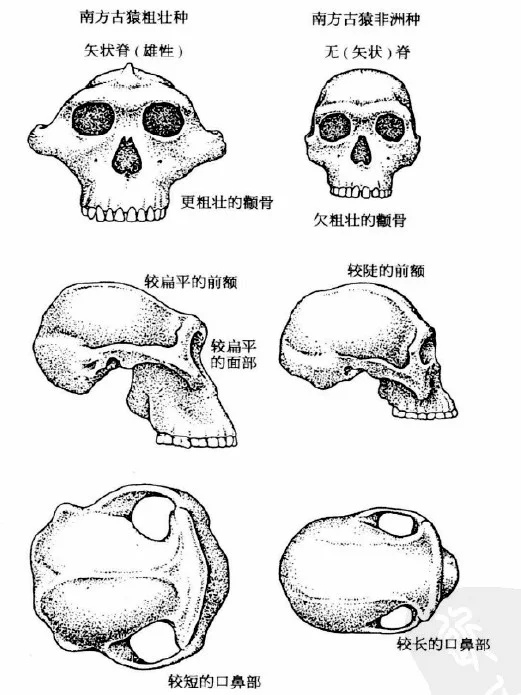
50年代人类学家们决定将已有的过多的人类的物种加以归并,只承认两个物种。当然,两者都是两足行走的猿,两者都是像汤恩小孩那样的似猿动物。两个物种的主要差别在于其颌骨和牙齿:两者的颌骨和牙齿都较大,但是一个物种比另一个物种更为粗壮(见图2-1)。较纤细的种被命名为南方古猿非洲种(Australopithecus africanus),这是达特1924年给予汤恩小孩的名称,这个名称的意思是“非洲的南方猿”。较粗壮的那个种被恰如其分地称作南方古猿粗壮种(Australopithecus robustus)。
1700189151
1700189152
1700189153
1700189154
1700189155
图2-1 南方古猿表兄弟 南方古猿粗壮种(和鲍氏种)与非洲种之间的主要区别在于咀嚼机制,包括颌骨的构造,颧骨和有关的肌肉附着处。粗壮种适应于包括粗糙的植物性食物,需要有力的咀嚼
1700189156
1700189157
从他们的牙齿构造上可明显地看出,无论非洲种或是粗壮种都是主要吃植物性食物的。他们的颊齿不像猿,后者有尖锐的齿尖,适合于吃相对较软的果子和其他植物。而南方古猿的颊齿却有平的研磨面。我设想,人类中最初的物种,如果以与猿相似的食物为生,他们会具有似猿的牙齿。在20O万年前到300万年前之间,人类的食物已明显地转变为比较坚硬的食物,诸如硬的水果和硬壳果,这几乎表明,南方古猿生活在比猿更为干燥的环境中。粗壮种的臼齿之硕大,暗示其所吃的食物特别坚硬,需要大面积的研磨。这些牙齿被归为“磨石臼齿”,不是没有原因的。
1700189158
1700189159
东非第一件早期人类化石是玛丽•利基于1959年8月发现的。在臭杜韦峡谷寻找堆积物几近30年后,她得到了报偿,发现了与南非的南方古猿粗壮种相像的磨石齿。可是奥杜韦的这个标本甚至比其南非的堂兄弟还要粗壮。与玛丽一起参加这项长期搜寻工作的路易斯•利基将这个标本命名为东非人鲍氏种(Zinjanthropus boisei)。属名意思是“东非的人”,boisei这个种名来自查尔斯•鲍伊斯(Charles Boise),他曾支持过我的父亲和母亲在奥杜韦峡谷和其他地方的工作。当现代地质测年方法首次应用于人类学时,东非人被确定为生活于175万年前。东非人这个名字最后变为南方古猿鲍氏种,因为它被假定为南方古猿粗壮种的东非变体或地理变异。
1700189160
1700189161
名称本身并不特别重要。重要的是我们正见到几个属于人类的物种,它们有着同样的基本适应性,两足行走,小的脑子和相对大的颊齿。这正是1969年我第一次到特卡纳湖东岸考察时在一条干涸的河床上发现的头骨上所看到的。
1700189162
1700189163
我们从各种骨头的大小了解到南方古猿的雄性比雌性大得多。雄性身高超过1.5米,而雌性几乎不到1.22米。雄性的体重几乎必定为雌性的两倍,这种差别今天我们可以在稀树草原狒狒中看到。因此可以合理地推测,南方古猿的社会结构与狒狒相似,正如上一章中已特别提到的,占优势的各个雄狒狒互相竞争,以接近成年的雌性。
1700189164
1700189165
发现东非人1年以后,我的哥哥乔纳森(Jonathan)也是在奥杜韦峡谷发现了另一种类型人类的头骨片,使得人类史前时代的故事变得稍稍复杂了。这块头骨片相对地较薄,表明这个个体比已知的各种南方古猿身体结构轻巧。它的颊齿较小,最重要的是,它的脑子几乎大出50%。我的父亲下结论说,虽然南方古猿是人类祖先的一部分,但这个新的标本代表最终产生现代人的那一支。在同行们一片喧嚣的反对声中,他决定将它命名为能人(Homo habilis),作为这个属的第一个早期成员(Homo habilis这个名字的意思是“手巧的人”,是达特向我父亲建议的,指的是设想这个物种是能制造工具的人)。
1700189166
1700189167
同行的喧嚣声有一些微妙的原因。其中部分原因是路易斯为了把人属这个名称用于这个刚被发现的化石,不得不把这个属的已被接受的定义加以改变。那时英国人类学家阿瑟•基思(Arthur Keith)爵士提出的人属的标准定义,包括人属的脑量应该等于或超过750毫升,这个数字介于现代人和猿之间,被认为是人和猿的脑量的界河。尽管在奥杜韦新发现的化石的脑量仅仅为650毫升,路易斯仍判断它属于人类,那是因为它的头骨更像人。因此他提出把人和猿的脑量的界河移到600毫升,从而让这个新的奥杜韦标本进入人属。这个战术无疑提高了随之而来的激烈争论的情绪。可是,这个新的定义最终还是被接受了(后来发展的结果是650毫升对能人的平均成年脑量来说是相当小的;800毫升才是一个接近的数字)。
1700189168
1700189169
除了科学名称外,还有一点很重要,就是由这些发现而开始形成的进化型式,即早期人类具有两种基本的类型。一种类型具有小的脑子和大的颊齿(这是各种南方古猿所具有的);另一种类型具有增大的脑子和较小的牙齿(人属)(见图2-2)。两者都是两足行走,但是在人属的进化中曾经明显地发生了某种十分异常的事件。在下一章中我们将更充分地探讨这个“某种事件”。无论怎么说,人类学家们对人类历史这一段(即距今200万年前后)的人的系统树的形状的了解,是颇为简单的。这树有两根主支:各种南方古猿,都在100万年前绝灭了;人属,最后导向像我们今天这样的人。
1700189170
1700189171

1700189172
1700189173