1700260100
要解决这样一个悖论,最好的办法就是“跳出来”。只要彻底跳出中心法则,不依赖碱基互补配对的手段获取RNA,一切问题就都迎刃而解了。所以说来说去,我们就是要讨论RNA的单体在没有模板和聚合酶的条件下,能不能聚合成链状的RNA。
1700260101
1700260102
如果认为只要有了RNA的单体,RNA就会在随机运动里自然而然地涌现出来,那就未免步了原始有机汤假说的后尘:稳定的溶液中不只有核苷酸聚合成RNA的反应,还有RNA水解成核苷酸的反应,而且后者要快得多,所以从化学平衡上看,RNA单体不会自发聚合成链,链状的RNA倒是会自发地水解成单体。
1700260103
1700260104
这个问题普遍存在,今天也同样不得不面对。例如,细胞里的RNA聚合酶负责按照DNA的模板链,用游离的核苷酸聚合出一条镜像序列的RNA,所以,它在理想情况下应该按着箭头指示的方向运动。
1700260105
1700260106
但是,在那个反应发生的位置,刚刚聚合的RNA也同样有可能发生水解反应,重新断裂成核苷酸[9],而且概率更大,如果不加干涉,RNA聚合酶其实会逆着箭头走——怎么办呢?
1700260107
1700260108
生命的解决之道当然就是能量:那些核苷酸同时是能量通货,细胞内的种种能量代谢会源源不断地制造它们,使它们总能维持非常高的浓度。这就让核苷酸漂进并参与反应的概率远远超过了漂出的概率,彻底扭转了那个化学平衡,让聚合酶总能顺着箭头前进了。
1700260109
1700260110
显然,在生命出现之前,地球上绝不可能有如此高效的核苷酸生成机制,也就绝不可能像细胞这样直接扭转化学平衡。这就是为什么在原始有机汤假说之后,一大批假说都将生命起源寄托在了陨石坑、火山口湖和火山温泉中——这些地方水量很少,在较高的温度下很容易发生周期性的干涸,其中的核苷酸也就能够周期性地浓缩起来,周期性地逆转那种化学平衡了。而热液喷口假说最常遭遇的一种质疑,就是这种环境位于深海,似乎很难把核苷酸持续地浓缩起来。
1700260111
1700260112

1700260113
1700260114
1700260115
图4—7 细胞内的RNA聚合酶工作原理示意图。这张图的样式和配色与这本书里其他的图明显不同,因为这张图是作者在写这本书之前画的。
1700260116
1700260117
然而这种质疑严重地低估了白烟囱假说。
1700260118
1700260119
白烟囱最独特的性质,就是它的内部充满了迷宫一样的毛细管道——仔细观察图3—11,你会看到那些沉积物就像疏松的海绵一样。超过40℃的热液和只有4℃的海水在其中缓慢流动着,形成了显著的温度差异,也就产生了强烈的“热泳效应”。
1700260120
1700260121
所谓“热泳效应”,是指在温度差异的作用下,介质中的微粒会在统计上向着温度较低的方向扩散。尤其对于那些住集中供暖的老房子的北方读者来说,这种效应听着陌生,却早已是生活中的烦恼了:暖气片比它紧邻的墙壁更热,所以空气中的灰尘微粒就会向着墙壁运动,天长日久,暖气片背后的那堵墙就会被“熏”得又黑又黄,怎么都洗刷不干净。
1700260122
1700260123
同样,分子较大的有机物如果出现在了白烟囱的毛细管道里,也会被热泳效应驱赶到温度较低的地方,在那里浓缩聚集起来。而且越大的有机物,越容易浓缩:核苷酸这种尺寸的分子可以通过这种机制浓缩至原本的1/5 000V,而当它们在这浓缩的过程中发生了聚合反应,浓缩的程度还会急剧增长。几个碱基长度的短RNA可以浓缩至原来的几十万分之一,上百个碱基的RNA链就可以浓缩至原来的数千亿分之一,而只需5厘米长的微管和10℃的温差,RNA就有可能聚合到200个碱基的长度VI。这实在是令人振奋的数据,要知道,真核细胞的转运RNA,也只有最多90个碱基的长度。
1700260124
1700260125
总之,在温度差异的作用下,核苷酸聚合与RNA水解的化学平衡立竿见影地扭转了,第一批RNA链也就应运而生了——换句话说,这位化学平衡·热泳先生,就是住在白烟囱里的RNA世界的大诗人,正是他,笔耕不辍地用RNA单体谱写出了一条条长链状的RNA分子。
1700260126
1700260127
不难想象,这位大作家颇有些率性,他只能把RNA单体随机地组合成长链,当然没有编码任何有意义的东西。但好在他也是个极有毅力的高产作家,对于长度只有几百个碱基的RNA,他可以在成千上万年的光阴中浓缩出无数条来,其中必然有一些具有独特的催化活性,能够以另一条RNA为模板,复制新的RNA出来。
1700260128
1700260129
这样的RNA一旦出现,中心法则的箭头就会开始运转,经典意义上的进化就会徐徐启动,一个全新的时代就此开始——但是,我们要把这些全都留在之后的章节里慢慢讨论,眼下趁这一章尚未结束,再提起另一个重要的伏笔。
1700260130
1700260131
你还记得第五章和第三幕里都出现过一些奇奇怪怪的辅酶吗?什么辅酶A、辅酶NADH、辅酶FAD、辅酶F420……统统是性质非常活跃的物质(参见图2—13或图3—2和图3—6),它们在各种化学反应之间运送各种官能团,让所有的生化反应交织成了错综复杂的网络。
1700260132
1700260133
如果仔细观察一下化学结构,我们就会发现它们都与RNA脱不开关系,其中一些最重要的辅酶根本就是核苷酸与其他物质杂糅出来的变体——第九章的第一篇“延伸阅读”还介绍过一种含有钴咕啉的辅酶,不妨再留神观察一下图3—16,那东西从下方勾住钴原子的,也正是一个腺苷酸的类似物。
1700260134
1700260135
考虑到这些辅酶广泛出现在一切生命的细胞内,显然也是末祖的古老遗产,所以RNA世界假说的研究者从一开始就在分析其中的曲折,他们推测这些辅酶很可能就是RNA世界的遗迹VII。
1700260136
1700260137
那位化学平衡·热泳先生既然是个随心所欲的家伙,他的连笔字想来也不工整。在最初的RNA世界,核苷酸的聚合反应并没有酶来催化,磷酸与核糖也就不像今天这样总能连成一条整整齐齐的骨架,而很可能一不小心就连出什么乱七八糟的结构,甚至与其他莫名其妙的物质杂糅起来。其中一些活性非常高,能够携带各种官能团的,就是原始的辅酶了。
1700260138
1700260139
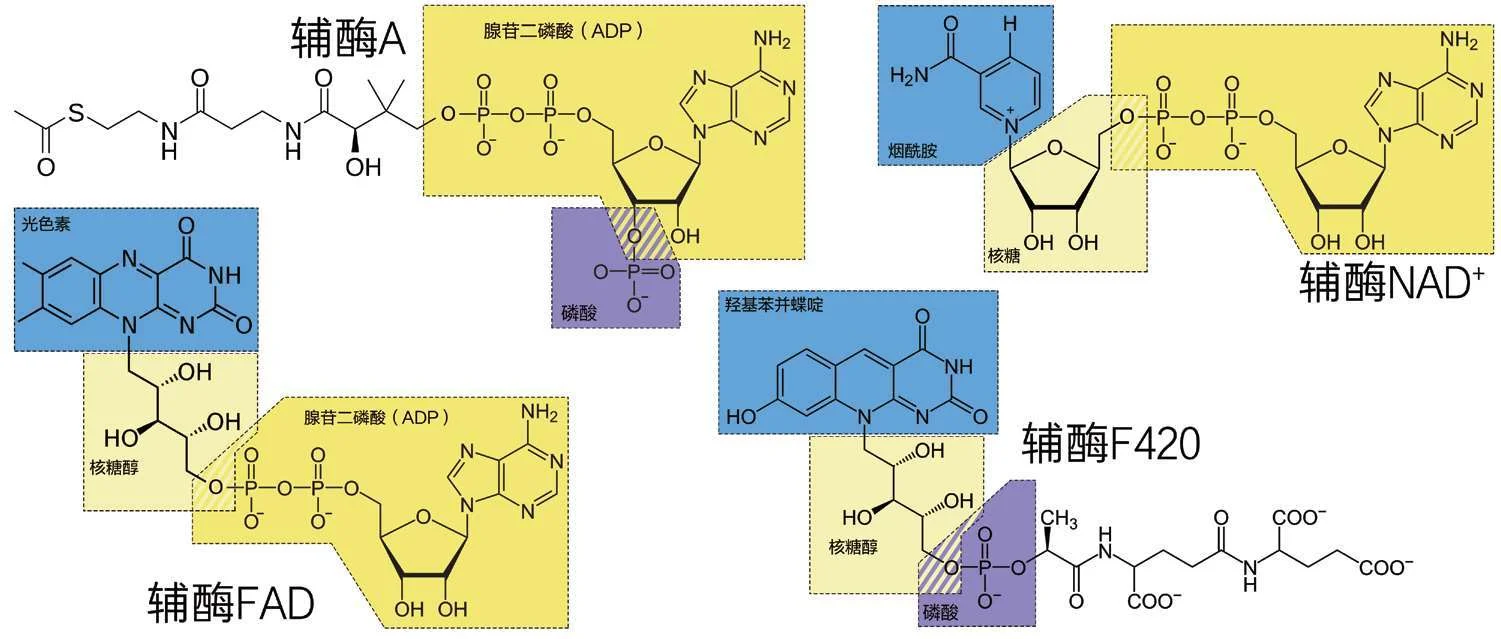
1700260140
1700260141
1700260142
图4—8 乙酰辅酶A路径和逆三羧酸循环里出现的4种重要的辅酶,它们在结构上都与RNA有些关系:深黄色的部分就是一个RNA单体,淡黄色的部分是核糖或者核糖醇,蓝色部分是标准碱基之外的几种含氮碱基。(作者绘)
1700260143
1700260144
想想看吧,这真是激动人心的想法:RNA世界才刚刚开始,我们就已经有了能量通货,有了各种辅酶,那么第三幕里的那些路径啊,循环啊,就与如今的细胞里的情形没什么不同了,这显然会极大地促进那些物质代谢与能量代谢,让一切运转都迅速起来——要是再能有些酶,那就完美了。
1700260145
1700260146
这可未必是个遗憾,因为一些锐意的研究者已经提出,这些辅酶就曾经是某些长链RNA的活性末端,因为这个位置上的核苷酸最容易杂糅其他物质,长链RNA也将因此成为强大的酶,实实在在地催化那些代谢反应。但在后来的进化中,蛋白质出现了,它们的催化能力更加强大,在进化中逐渐取代了RNA的长链部分,只把RNA的活性末端保留下来,那活性末端就以辅酶的身份在进化中继续发挥作用,直到今天VIII。
1700260147
1700260148

1700260149
[1]“甲醛聚糖”的英文作“formose”,乍看上去很像拉丁语的“美丽”(formosa),但它们没有任何词源上的关系。“formose”是截取了“formaldehyde”(甲醛)的前半部分和“aldose”(醛糖)的后半部分,“混成”来的。在这里,“混成”是一种在英语里很常见,在汉语里却十分罕见的构词法,如果要“信、达、雅”地直译一个混成词,就会非常棘手,所以“甲醛聚糖”这四个字就完全是意译来的。但是,如果一定要追求优雅的直译,我们又会得到一些妙趣横生的故事——1968年,中国的汉语言学之父赵元任先生翻译了路易斯·卡罗尔的《爱丽丝镜中奇遇记》,在处理那首英语文学里最杰出的荒诞诗《炸脖伏诛记》的时候,就遇上了一连串的这样的难题。然而这些难题、那首诗的形式,以及那首诗的内容,都刚好与这一章的内容充满了有趣的联系。实际上,这本书的这一章原本并不是这样写成的,这本书的作者试着重新翻译了那首诗,讲述一个小英雄屠龙与RNA世界极早期交错起来的故事。出版社的编辑为我这个精妙的奇想掉了太多头发,为了保住他们剩余的秀发,这本书的作者不得不删掉了他很喜欢的大部分内容,只留下你现在看到的这一部分。不过,读到这里,你仍然有机会看到那原来的写法,因为这本书的作者也把原本的这一章发表在了他的微博上,你可以通过下面的网址,找到这一章完整的样子。