1700258692
1700258693
而生命用来组建蛋白质的氨基酸多达22种[23],一个蛋白质又可以含有成千上万个氨基酸,由此产生的序列差异可谓无穷,这也就从根本上决定了蛋白质拥有几近无穷的多样性。
1700258694
1700258695
如果仅仅是一条氨基酸缩合成的长链,那就只能叫作“多肽”而已。但是请注意,氨基和羧基缩合成的肽键是个好东西:它残余了一个连在氮上的氢,这个氢带有很强的正电荷;还有一个连在碳上的氧,这个氧带有很强的负电荷。那么,在整个多肽长链上,不同部位的这两种原子就会强烈地吸引,形成很强的氢键[24]。于是,一条很长的多肽就会因为这些氢键而不断弯曲,出现螺旋、片层和拐角等“二级结构”。
1700258696
1700258697
不仅如此,我们还要注意到,那20种氨基酸每一种都有一个独特的侧链(R),这些侧链各自具有独特的化学性质,还与水有各不相同的亲和能力,而细胞内哪哪都是水,多肽存在于这样的环境里,就会被迫发生一些形态变化。
1700258698
1700258699
水分子会把那些亲水的氨基酸向外拉扯,同时把那些憎水的氨基酸向内推挤。多肽于是皱缩起来,折叠成团,那些侧链也趁机发生各种相互作用,进一步改变这团多肽的三维结构。
1700258700
1700258701
上面这整个过程听起来很混乱,让人感觉一条多肽链经过这么一折腾,就成了一团乱麻,但事情完全不是这样。麻绳随手一团会变得乱七八糟,是因为它是个未加限制的宏观物体,可以在任何一点向任何方向弯曲。但在分子的世界里,化学键是量子化的,也就是说,化学键不管怎么摇摆、伸缩、转动,相互之间必须满足特定的空间关系。而肽键尤其讲究,它附近的4个化学键(图2—71虚线内的6个原子构成的4个化学键)必须在同一个平面内小范围地运动[25]。所以,多肽链虽叫“链”,却不像串珠项链那样想怎么盘就怎么盘,而更像自行车的链条,只能在非常有限的范围之内变化。
1700258702
1700258703

1700258704
1700258705
1700258706
图2—71 肽链上的4个氨基酸的片段。绿色的键,就是肽键。注意,红色的是连接在碳上的氧,蓝色的氮上有一个氢,氢和氧总是处在相反的两侧。(作者绘)
1700258707
1700258708

1700258709
1700258710
1700258711
图2—72 正中间的是“胱天蛋白酶3号”(Caspase 3),这种蛋白质与细胞的程序性死亡关系密切,与癌症和衰老息息相关,但我们只关心它的二级结构:涂成红色的是α螺旋,涂成蓝色的是β片层,左右两边分别放大了它们。可见,是肽键之间的氢键,也就是那些断珠线,将细长的肽链约束成了这些具体的形态。(作者绘)
1700258712
1700258713
但是,自行车链条除了相邻的两节会彼此铰接,其余任何两节之间就都再没有什么相互作用了,而多肽链却不一样。那些氨基酸残基之间存在着各种相互作用,就如同这链条的某些节上长了钩子,某些节上长了扣子,某些节上长了搭子,某些节上长了套子……如图2—73,这些小结构互相匹配,非要盘成某个特定的形状才能稳定。而经过疏水作用和氨基酸残基的相互作用,多肽最后盘成的特定形状,就被称为蛋白质的三级结构。
1700258714
1700258715
可是,这成千上万的氨基酸残基互相匹配,就不会乱套吗?
1700258716
1700258717
可能性当然有,但实际上很难发生,因为多肽链并不会同时暴露所有的残基,一窝蜂地匹配,而是从多肽链的一段开始,一个残基一个残基地匹配。
1700258718
1700258719
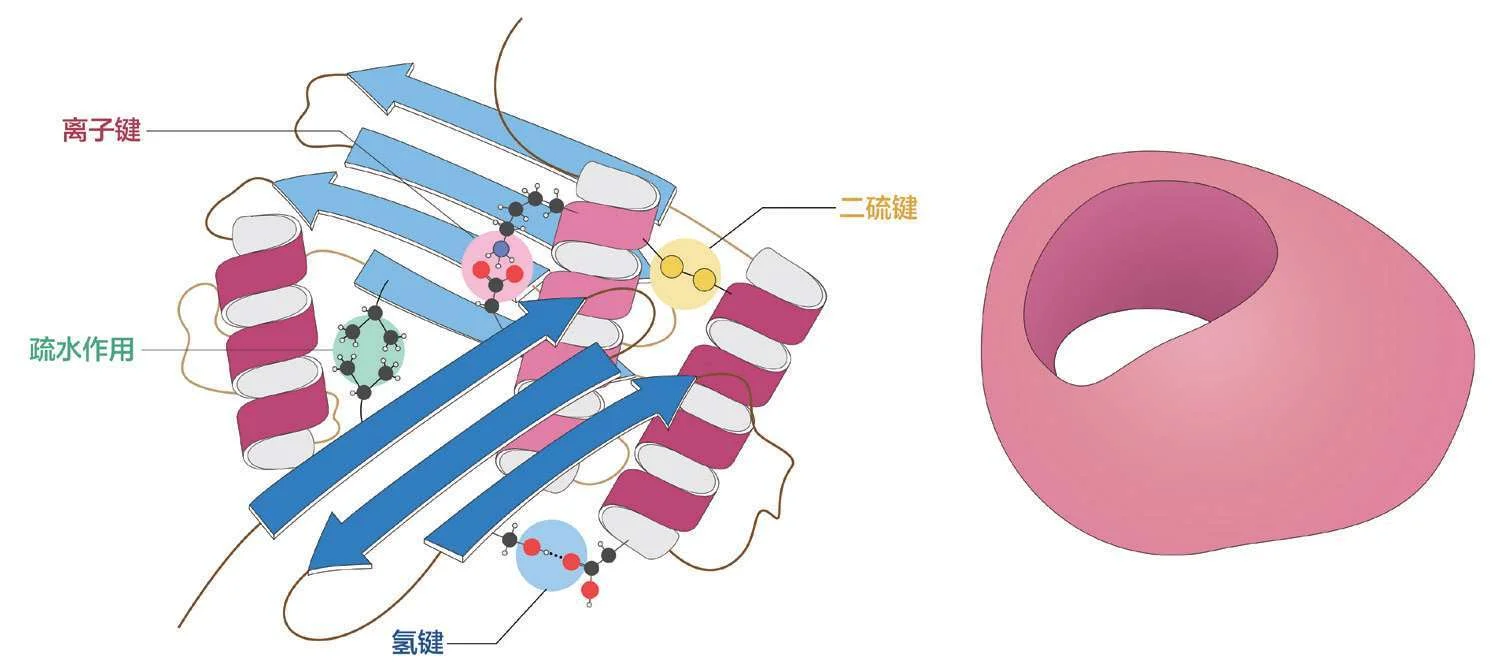
1700258720
1700258721
1700258722
图2—73 这是一个虚构的蛋白质,只标注了影响三级结构的氨基酸残基,它们彼此之间的相互作用让二级结构中的螺旋和片层进一步组织成某种三级结构——近似一个带有大窟窿的怪异球体。(作者绘)
1700258723
1700258724
在前文中,我们先认识了蛋白质的一级结构,再认识了二级结构,最后才认识了三级结构,所以难免会产生一种误解,以为蛋白质是先合成一条完整的多肽链出来,然后再想办法把这漫长的链条,按照二级结构和三级结构的要求,折叠成型。
1700258725
1700258726
但事情不是这样的。氨基酸在核糖体上缩合成肽链,这条肽链从核糖体的出口一露出来,就会立刻开始折叠。在基本化学规律的作用下,第一节和第二节先折叠成稳定的三维形态,第三节再和前两节折叠成稳定的三维形态,第四节再和前三节折叠成稳定的三维形态,以此类推。
1700258727
1700258728
所以,那些二级结构和三级结构并非折叠的“要求”,而是折叠的“动力”。刚露出来的残基和之前露出来的残基凭借氢键强烈吸引,就会不可避免地扭曲成螺旋,弯曲成片层,因此呈现出了二级结构。在此基础上,氨基酸的残基又不可避免地凭借其他作用相互吸引,形成更加复杂的三级结构,而不是反之为了获得预定的二三级结构,要用某种力量把肽链拧成想要的螺旋,再捆上各种化学作用使其就范。因此,在确定的温度下,多肽链的一级结构是确定的,折叠的过程就总是确定的,二级结构和三级结构也就总是确定的。
1700258729
1700258730
如果仍要打个比方,氨基酸在核糖体上缩合成多肽链并折叠出初步的三维形态,这个过程就好像一个人在织毛衣,他只能从毛线团上抻一点线,织几针,再抻一点,再织几针,随抻随织,该怎么织就怎么织,而不是把几百米长的毛线一次性全抖搂开,再整个人扑进去“兴风作浪”。
1700258731
1700258732
总之,以这种小心翼翼,边合成边折叠的方法,绝大多数的蛋白质一经合成就已经折叠完善,可以行使正常的生化功能了。
1700258733
1700258734
但另外那些蛋白质该怎么办呢?
1700258735
1700258736
也不难办。它们虽然还不足以行使预期的生化功能,但也已经有了独一无二的形态,可以被其他蛋白质识别了。在这些“其他蛋白质”中,有的能帮助没折叠好的蛋白质折叠好,达成预期的三维形态,比如营造小范围的疏水环境让部分结构从里往外翻转[26];有的能催化氨基酸的侧链形成特殊的化学键,构成特殊的三维结构,比如帮助角蛋白形成大量的二硫键,缠结得致密坚硬,用来生长指甲和毛发;有的能给蛋白质插入其他构件,比如给血红蛋白加载血红素,给复合物I镶嵌铁硫簇;还有的是几个蛋白质能够互相识别,像乐高积木一样组装起来,形成更大更复杂的四级结构,比如电子传递链上的各种复合物、水轮机似的ATP合酶以及核糖体,都是拥有四级结构的复合物。
1700258737
1700258738
总而言之,只要信使RNA上的碱基序列是确定的,核糖体缩合多肽时的氨基酸序列就是确定的,蛋白质的三维形态就是确定的,它的生化功能就是确定的,产生的生理作用就是确定的,一切都是相当确定的。[27]
1700258739
1700258740
最后,当然,我们还是要说得严谨一些,上一段的“确定”也不是绝对的。某些蛋白质实在折叠不好也是常事。此时,它们通常就会被某些“回收蛋白”识别出来,及时销毁,水解成氨基酸拿去制造新的蛋白质。而在极端情况下,某些蛋白质既不能正常折叠也不能回收,就会招致大麻烦——阿尔茨海默病、帕金森病和疯牛病,都与蛋白质不能正常折叠也不能正常回收有关。
1700258741