1700261615
生命的起源:所有生命的共同祖先在40亿年前是怎样诞生的? [:1700256355]
1700261616
·螺旋与发卡·
1700261617
1700261618
早在第六章我们就说过,核糖体中既有RNA,也有蛋白质,但RNA的地位更重要。又如第十三章强调过的,2000年,我们确定了核糖体RNA也是一种酶RNA,是它催化了从氨基酸到肽链的聚合反应,而蛋白质在此过程中只起强化结构的作用。这也是RNA世界假说的主要依据之一。
1700261619
1700261620
但核糖体RNA实在是太难观察,太难研究了,它一方面是已知最大的分子结构,复杂性超越一切蛋白质或核酸的复合物,另一方面它又仍然很微小:大亚基的直径只有20纳米到30纳米,放大三五十倍都能轻松穿过医用外科口罩的过滤层,其中的RNA与蛋白质更是纠缠不清,好像沙滩上拧成团的破渔网,而那个关键的聚合反应,却只发生在核心深处千分之几的体积内,一个被称为“肽基转移酶中心”的地方,难窥究竟。
1700261621
1700261622
为此,从20世纪70年代开始,以色列的晶体生物学家阿达·约纳特做了30多年的持续研究,终于弄明白了肽基转移酶中心的精确结构和催化原理I,荣膺2009年的诺贝尔化学奖,也成为有史以来第一个获得诺贝尔奖的亚洲女性。
1700261623
1700261624
约纳特的研究揭示了这样一个耐人寻味的事实:肽基转移酶中心是一对180°对称的螺旋,它们以不可思议的精度形成了一个对称的漏斗,迫使氨基酸在狭窄的空间里缩合成肽链。这个对称的漏斗就如图4—70所示,你不妨先花一些时间仔细看看它,找找其中的螺旋。
1700261625
1700261626

1700261627
1700261628
1700261629
图4—68 左侧的是细菌核糖体的小亚基,浅绿色的是RNA,深绿色的是蛋白质,红色的是1分子四环素,它能强占信使RNA的结合位点,使细菌因无法制造蛋白质而死亡;右侧的是细菌核糖体的大亚基,浅蓝色的是RNA,深蓝色的是蛋白质,红色的是1分子氯霉素,它能霸占氨基酸聚合反应的核心,使细菌因无法制造蛋白质而死亡。(DavidGoodsell绘)
1700261630
1700261631

1700261632
1700261633
1700261634
图4—69 2009年的诺贝尔化学奖颁给了文卡特拉曼·拉马克里希南(左)、托马斯·施泰茨(中)和阿达·约纳特(右),嘉奖他们“对核糖体结构与功能的研究”。(来自Royal Society uploader、Prolineserver、Science History Institute)
1700261635
1700261636
1700261637
1700261638
1700261639
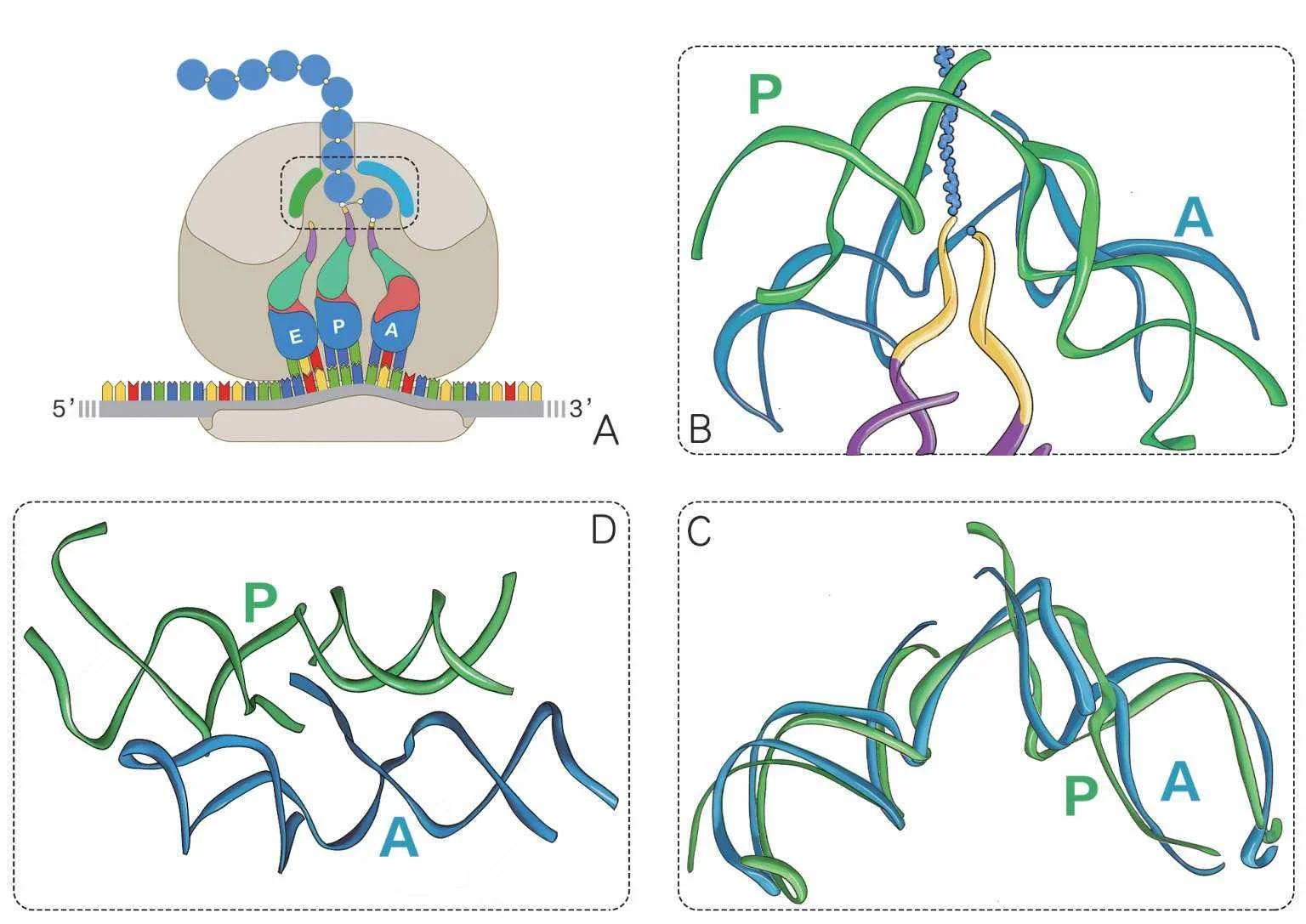
图4—70 肽基转移酶中心的结构。A图像就是第六章里,我们温习中心法则的翻译过程的图2—51,这里额外标注了一段蓝色和绿色的弧线,那就是肽基转移酶中心在核糖体中的位置,它们约束了P位点和A位点两个转运RNA的末端,促成了肽键的形成。更真实的B图像来自阿达·约纳特在2009年的诺贝尔奖演讲稿中使用的插图,上方的蓝色螺旋和绿色螺旋就是那两段弧线的真实形态。这是两个高度相似的RNA螺旋,图中只画了它们的磷酸核糖骨架,省略了所有的碱基,可见二者共同围绕成了一个倒置的漏斗状,下方黄色的是两个转运RNA的CCA尾,它们伸进了漏斗的中心。如果你没有从这个角度看出两个螺旋的相似与对称,那么C图像是把P螺旋旋转180°的结果,你会看到它与A螺旋有着惊人的重合。D图像是从下方仰视这个漏斗的样子,你会看到这两个螺旋相似且对称。(作者绘)
1700261640
1700261641

1700261642
1700261643
1700261644
图4—71 如果上面的讲述还是不太能让你理解这整个过程,那么把图4—70中的D图像抽象一下,就得到了这张图。蓝色和绿色的螺旋分别是肽基转移酶中心的A螺旋和P螺旋,标记着“A”和“P”的倒锥形复合物就是A位点和P位点的两个转运RNA的末端(因为是仰视图,所以它们都截断了,用虚线表示截面)。那么,你会看到A和P两个螺旋的凹槽刚好对在一起,两个转运RNA的末端刚好沿着这个凹槽滑动。同时,凹槽底部正中央是漏斗的出口,P位点的转运RNA携带的肽链就从这个出口顺出去,而A位点的转运RNA带着一个新的氨基酸从右下方滑进凹槽,当它足够靠近肽链的末端,整个肽链就会连接到这个新的氨基酸上面了,即所谓的“肽基转移”——这整个过程对应的,就是图2—51里发生的变化。(作者绘)
1700261645
1700261646
结合第六章,我们会发现,在具体的翻译过程中,相继进入核糖体的两个转运RNA都会把CCA尾伸进漏斗,其中,先来的CCA尾上是已经形成的肽链,后到的CCA尾上是下一个氨基酸。而在漏斗收拢的中心,这个氨基酸就会与肽链靠得足够近,由此形成又一个肽键,让整条肽链延长一节。这个结构非常精密,两个螺旋稍微改变一个碱基都可能改变蛋白质的合成效率。
1700261647
1700261648
所以迄今为止,人类至少比较了三个域的930个物种,连同线粒体和叶绿体的这对螺旋,结果发现它们的肽基转移酶中心都惊人地一致,一致得可以重叠起来。循着我们在这一幕里的“经验”,我们已经清晰地感觉到,这又是末祖的遗传。于是,约纳特在2009年的诺贝尔奖颁奖现场的演讲稿中提出:核糖体最初的形态就是这两个对称的螺旋,除此之外层层叠叠的RNA与蛋白质,全都是在进化中逐渐附加上去的。
1700261649
1700261650
生命的起源:所有生命的共同祖先在40亿年前是怎样诞生的? [:1700256356]
1700261651
·更多的螺旋·
1700261652
1700261653
如果这里不断提起的“螺旋”二字让你回想起上一章的某些内容,想到了转运RNA的迷你螺旋,那你的确想到关键地方去了:如图4—72,肽基转移酶中心的两个螺旋,竟然与转运RNA如此相像。II
1700261654
1700261655

1700261656
1700261657
1700261658
图4—72 蓝色和绿色的是肽基转移酶中心的两个螺旋,橙色的是转运RNA,它们的三维结构可以很近似地重叠起来。(作者绘)
1700261659
1700261660
最早注意到这种相似性的,是另一个优秀的以色列女性化学家伊拉纳·阿格蒙(Ilana Agmon)。她与阿达·约纳特一同在魏茨曼科学研究所[1]工作,是阿达·约纳特的研究小组里的重要成员,许多关于核糖体结构的突破性成果都凝聚着她的夙寐辛劳。在过去的10余年里,阿格蒙不但发现了图4—72里那种微妙的相似性,还在核糖体里注意到了另外一些蹊跷之处。
1700261661
1700261662
想想看,我们已经有了转运RNA,也有了肽基转移酶中心,但要真的实现翻译过程,这还远远不够。因为如果只有它们,肽基转移酶中心就只能随机结合转运RNA,聚合出随机序列的肽链——当然,随机的肽链也未必完全无用,我们会在下一幕里看到一些关于它们的消息,但毕竟,我们期待的是真正的蛋白质翻译系统,所以还必须有别的东西能让信使RNA也参与进来,给转运RNA赋予次序,还要有什么东西能把这一切固定在恰到好处的空间位置上,这样才能成为一个实在的蛋白质翻译系统。
1700261663
1700261664
在今天的核糖体里,许多RNA或蛋白质部件都参与了这两项任务。不过追究起来,负责结合信使RNA的关键结构是小亚基的“3’端附近结构”,把这一切固定起来的关键结构是大亚基里的“桥元件”。[2]在今天的细胞里,它们的位置关系就像如图4—73展示的那样。