1700258400
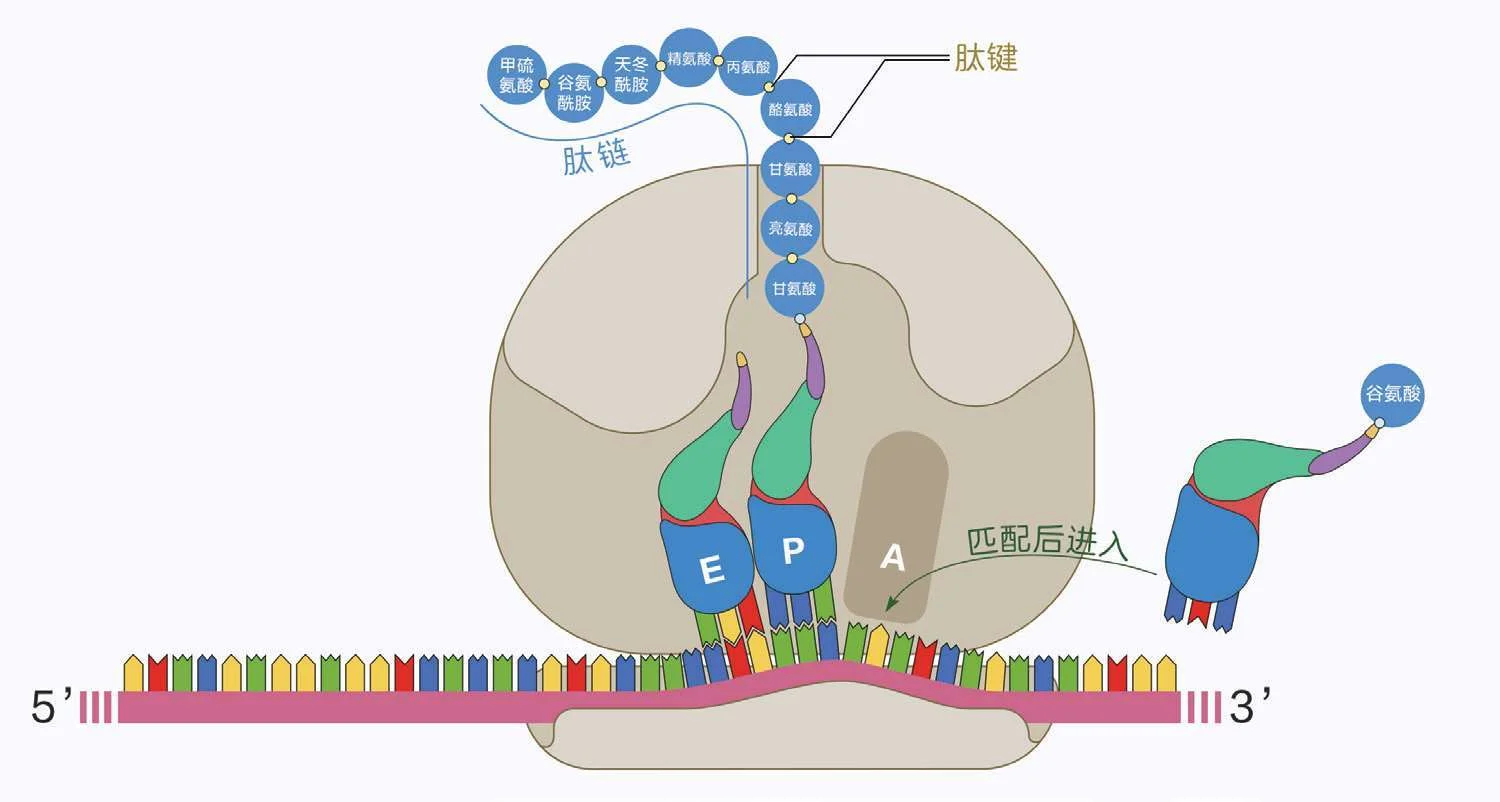
信使RNA与核糖体装配完毕,细胞质里不计其数的转运RNA就要开始工作了。如图2—49,转运RNA的三维形状一头粗来一头细,中间还拐个弯,好像一只鸡大腿,大腿尖上粘着一个氨基酸,大腿根向外翻出3个碱基,而且这3个碱基一定是那个氨基酸的密码子的互补序列。比如大腿尖上粘的是谷氨酸,谷氨酸的密码子是“GAG”或者“GAA”,那么大腿根上翻出来的3个碱基就一定是“CUC”或者“CUU”——像这样与密码子互补的3个碱基,就是所谓的“反密码子”。
1700258401
1700258402
这些转运RNA的工作室就在核糖体大亚基的内部,为此,这个大亚基有3个关键的出入口。
1700258403
1700258404
首先是“转运RNA入口”。如图2—49至图2—52,装载了氨基酸的转运RNA会一个一个地从这里钻进去,试着匹配信使RNA上的密码子。如果某个转运RNA匹配成功了,它就会结合在这个密码子上,钻进核糖体内部,到达“A”位点上;如果匹配不成功,那当然就得从入口退出来,让别的转运RNA来试。
1700258405
1700258406

1700258407
1700258408
1700258409
图2—49 核糖体的大小亚基组装在了信使RNA上。通过大亚基画成半透明的部分,可见其中同时预装了一个携带着甲硫氨酸的转运RNA,那是因为遗传密码的“起始密码”同时编码了甲硫氨酸(参见图2—44),所以新合成的肽链总是以甲硫氨酸开头的。至于那些不需要这个开头的肽链,就等肽链形成之后,再由别的酶来把它剪掉。(作者绘)
1700258410
1700258411
1700258412
1700258413
1700258414
图2—50 正在翻译中的核糖体,一个装载了氨基酸的转运RNA匹配后进入核糖体。为了表现清晰,大亚基展现的是内部结构。(作者绘)
1700258415
1700258416

1700258417
1700258418
1700258419
图2—51 核糖体内部的“肽基转移”。肽链原本结合在P位点的转运RNA上,现在要与A位点的转运RNA上装载的氨基酸形成新的肽键,由此整个转移到A位点的转运RNA上。那些仔细观察的读者会注意到A位点上的转运RNA的图示有些与众不同,因为这里试图表现的是这个转运RNA的“背面”:与所有酶促反应一样,这个图示里的一切发生在三维空间中,A位点和P位点的两个转运RNA的位置关系,就好像我们坐在凳子上把彼此的脚抵在一起。那么,此时低头俯视双脚,我们看到的就会是图示中的这个样子——只是我们的两条腿左右对称,所以低头看到的都是腿内侧,而转运RNA都是一顺子,所以一个看到正面,一个看到背面。另外,你还会注意到小亚基上的信使RNA不是平直的,而是在中间升起了一节,那就是专门给两个转运RNA预备的“凳子”。(作者绘)
1700258420
1700258421
在核糖体的深处,有一条正在形成的肽链,连接在P位点的转运RNA上。那么如图2—51,每一个到达A位点的鸡大腿都会朝它“磕”一下,由此把大腿尖上的氨基酸连到那个肽链的末端,形成一个新的肽键,让肽链再延长一节。这也让那条正在形成的肽链转移到了A位点的转运RNA上,因此,这个A位点的转运RNA就会继续深入,顶替掉原本的P位点的转运RNA。当然,原本的P位点的转运RNA也会顺势退让到E位点上,然后从另一侧的转运RNA出口离开核糖体。
1700258422
1700258423
这个过程循环往复,肽链就会不断延长,从上方的肽链出口钻出来,逐步折叠成细胞所需的一切蛋白质。
1700258424
1700258425
相比中学生物教材,上面的介绍似乎要更加精致复杂一些,因为我们会在第四幕里追究一些更加深刻的问题。但即便如此,这样的描述仍然省去了太多的细节,略去了太多的辅助物质。然而,我们实在不必纠结那些细节和物质,因为我们只需记住,在这整个过程中,信使RNA、核糖体RNA以及转运RNA,它们的结构,它们发挥功能的机制,它们彼此配合的方式,在如今的每一个细胞内都是一样的。当然,这种一样不是说没有差异的绝对相同,进化当然会让不同类群的RNA产生一些自己的特征,但这些特征都是修饰性的,丝毫不能掩盖它们源自同一套原型的昭然事实,这就像不同的鸟有不同形状的喙和不同长度的腿,但其中的相似性也实在显著,毫无疑问是来自同一套原型。
1700258426
1700258427

1700258428
1700258429
1700258430
图2—52 翻译过程中一个循环结束的样子。仔细看,这其实与图2—50一模一样,只是整个核糖体向右前进了一个密码子,肽链也增加了一个氨基酸。(作者绘)
1700258431
1700258432
我们发现,遗传密码、信使RNA、核糖体RNA以及转运RNA,乃至整个中心法则,都必然源自末祖,也就很可能源自第一个细胞——这毫无疑问就是我们在回溯之路上的又一条具体线索,恐怕也是最醒目的线索。
1700258433
1700258434
所以我们已经看到,这条线索吸引了最广泛的注意,半个多世纪以来,任何一个科学的生命起源理论都倾注了大量的精力去寻找核酸与蛋白质的起源,构想它们二者是如何在40亿年前关联了起来,但是任何这样的构想都会不可避免地遇上一个恼人的难题。
1700258435
1700258436
生命的起源:所有生命的共同祖先在40亿年前是怎样诞生的? [:1700256303]
1700258437
·先迈哪只脚?·
1700258438
1700258439
这个恼人的难题就是分子生物学版的“先有鸡还是先有蛋”:
1700258440
1700258441
最直接的问法是,核酸和蛋白质,到底谁先出现?
1700258442
1700258443
乍看起来,蛋白质的氨基酸序列全都预存在核酸的碱基序列中,所以是先有核酸再有蛋白质。可再一看,中心法则的每一个箭头,又都是在各种蛋白质的催化之下才得以实现的,尤其是DNA的复制,竟然需要几十种蛋白质来协调,所以看来是先有蛋白质,再有核酸。
1700258444
1700258445
更进一步,遗传和代谢,到底是谁先出现?
1700258446
1700258447
在这一章的前半部分,我们用生命的信息描述了一切生命活动,当然也包括一切代谢活动,而生命的信息归根结底全都来自遗传,所以看来是先有遗传,后有代谢。但是简述了中心法则之后,我们又发现,只有代谢活动已经完善并确立,能维持物质和能量的恰当供应,整套遗传活动才能周而复始地循环起来。所以,是先有代谢,后有遗传。
1700258448
1700258449
就这样,在探索生命起源的道路上,遗传和代谢究竟应该先迈哪只脚,似乎成了一个尴尬的死循环。